
 Home
Home
研究内容
当研究室では生体の力学的現象を扱うバイオメカニクス、とくに細胞に加わる力学的刺激とその刺激に対する細胞の感知・応答機構に焦点を当てた研究を行っている。研究の主題は血流に起因する力学的刺激であるシェアストレス(shear stress)とそれが作用する血管内皮細胞の関係を探ることである。このことは血流を介した血管系システムの制御機構の解明に役立つだけでなく、血流依存性に起こる血管の新生や成長やリモデリングあるいはヒトの粥状動脈硬化症や動脈瘤の発生といった臨床医学的に重要な問題の解明にも繋がる。
研究方法は培養した内皮細胞に流体力学的に設計した流れ負荷装置 で定量的なshear stressを作用させて細胞応答を観察する独自の生体工学的実験を用いている。これまで行ってきた研究の成果を以下の3項目に分けて紹介する。
で定量的なshear stressを作用させて細胞応答を観察する独自の生体工学的実験を用いている。これまで行ってきた研究の成果を以下の3項目に分けて紹介する。
1.Shear stressに対する細胞応答
2.Shear stressによる遺伝子発現制御機構
3.Shear stressの感知・情報伝達機構
Shear stress に対する内皮細胞応答
 Shear stress
Shear stress Static内皮細胞がshear stressに反応して多くの細胞機能を変化させることを明らかにした。例えば、培養内皮層に人工的に剥離部をつくると、周辺の内皮細胞が遊走・増殖して剥離部を修復するが、shear stressは内皮細胞の遊走・増殖を刺激し剥離部再生を促進した(Micorvasc Res 1987, Biorheology 1990)。また、shear stressが内皮細胞の一酸化窒素(NO)の産生を、shear stressの強さ依存性に亢進させること(BBRC 1994)、及び抗血栓活性を発揮するトロンボモデュリンの細胞膜発現量を増加させること(BBRC 1994)を観察した。併せて、shear stressが内皮細胞と白血球との接着に関わる接着分子VCAM-1(vascular cell adhesion mokecule-1)の細胞膜発現量を減少させ、リンパ球の接着を抑制することを示した(BBRC 1993, Am J Physiol 1994)。共同研究によりshear stressがNOと同様血管拡張作用を持つC-型利尿ペプチド、アドレノメデュリンのmRNA レベルを上昇させること(Hypertension 1997)、また、新しく発見された酸化型低比重リポ蛋白受容体の蛋白およびmRNA レベルを増加させること(Circ Res 1998)を明らかにした。最近、ヒトの末梢血を流れる内皮前駆細胞がshear stressに反応して増殖、分化、管腔形成能が亢進することを観察した(J Appl Physiol 2003)。また、shear stressがマウスの胚性幹細胞(ES細胞)を内皮細胞へ分化誘導する効果があることを明らかにした(Am J Physiol 2005)。このことを応用し、ポリマーの管にES細胞を播種し拍動性のshear stressを与えることで生体の血管に近い組織を持つハイブリッドの人工血管の開発を行うことができた(J Artif Organs 2005)。また、cyclic strain刺激がマウスのES細胞を平滑筋細胞へ分化誘導し、その効果に血小板由来増殖因子受容体のリガンド非依存性の燐酸化が関わっていることを明らかにした(J Appl Physiol 2008)。さらに最近、shear stressが内皮前駆細胞やマウスのES細胞において動脈内皮のマーカーであるephrinB2の発現を増加させる反面、静脈内皮マーカーであるEphB4の発現を抑制することから、shear stressが内皮の動静脈分化にも影響を及ぼすことを示した(J Appl Physiol 2009, Arterioscler Thromb Vasc Biol 2009)。
Static内皮細胞がshear stressに反応して多くの細胞機能を変化させることを明らかにした。例えば、培養内皮層に人工的に剥離部をつくると、周辺の内皮細胞が遊走・増殖して剥離部を修復するが、shear stressは内皮細胞の遊走・増殖を刺激し剥離部再生を促進した(Micorvasc Res 1987, Biorheology 1990)。また、shear stressが内皮細胞の一酸化窒素(NO)の産生を、shear stressの強さ依存性に亢進させること(BBRC 1994)、及び抗血栓活性を発揮するトロンボモデュリンの細胞膜発現量を増加させること(BBRC 1994)を観察した。併せて、shear stressが内皮細胞と白血球との接着に関わる接着分子VCAM-1(vascular cell adhesion mokecule-1)の細胞膜発現量を減少させ、リンパ球の接着を抑制することを示した(BBRC 1993, Am J Physiol 1994)。共同研究によりshear stressがNOと同様血管拡張作用を持つC-型利尿ペプチド、アドレノメデュリンのmRNA レベルを上昇させること(Hypertension 1997)、また、新しく発見された酸化型低比重リポ蛋白受容体の蛋白およびmRNA レベルを増加させること(Circ Res 1998)を明らかにした。最近、ヒトの末梢血を流れる内皮前駆細胞がshear stressに反応して増殖、分化、管腔形成能が亢進することを観察した(J Appl Physiol 2003)。また、shear stressがマウスの胚性幹細胞(ES細胞)を内皮細胞へ分化誘導する効果があることを明らかにした(Am J Physiol 2005)。このことを応用し、ポリマーの管にES細胞を播種し拍動性のshear stressを与えることで生体の血管に近い組織を持つハイブリッドの人工血管の開発を行うことができた(J Artif Organs 2005)。また、cyclic strain刺激がマウスのES細胞を平滑筋細胞へ分化誘導し、その効果に血小板由来増殖因子受容体のリガンド非依存性の燐酸化が関わっていることを明らかにした(J Appl Physiol 2008)。さらに最近、shear stressが内皮前駆細胞やマウスのES細胞において動脈内皮のマーカーであるephrinB2の発現を増加させる反面、静脈内皮マーカーであるEphB4の発現を抑制することから、shear stressが内皮の動静脈分化にも影響を及ぼすことを示した(J Appl Physiol 2009, Arterioscler Thromb Vasc Biol 2009)。
Shear stress による遺伝子発現制御機構
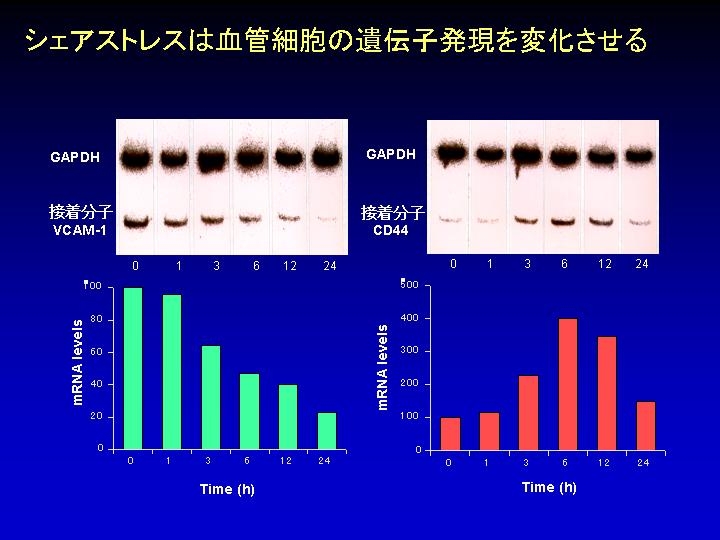 クリックすると拡大します。Shear stressが内皮細胞の遺伝子の発現を転写調節あるいは転写後調節することを明らかにした。転写調節に関しては、VCAM-1遺伝子の発現がshear stressで抑制を受けるが、それには遺伝子プロモータに2個並んで存在する転写因子AP-1結合エレメント(TGACTCA)がshear stress応答配列として働いていることを示した(Am J Physiol 1997)。転写後調節に関しては、顆粒級・マクロファージ・コロニー刺激因子(GM-CSF)の遺伝子発現がshear stressで増加するが、その効果は転写ではなくmRNAの安定化を介していることを明らかにした(Cric Res 1998)。また既知の遺伝子だけでなく多くの未知の遺伝子もshear stressに感受性のあることをmRNAのdifferential display法で示した(BBRC 1996)。共同研究によりshear stressに反応するG蛋白受容体ファミリーに属する未知の遺伝子をクローニングした(BBRC 1997)。また、流れで誘発されるCa2+反応に関わるP2X4プリノセプターの発現がshear stressで抑制を受けるが、これは転写因子SP1が関連した転写抑制に基づくことを明らかにした(Am J Physiol 2001)。さらにDNAマイクロアレイ解析で遺伝子全体の約3%(約600の遺伝子に相当)がshear stressに応答することを観察した(J Athero Thromb 2003)。最近、遺伝子に対するshear stressの作用が層流と乱流で異なることを明らかにした。線溶と血管のリモデリングに関わるウロキナーゼ型のプラスミノーゲン・アクチベータ(uPA)の遺伝子の発現は層流で低下し、乱流で増加した。層流は転写因子GATA6を活性化し転写を抑制するとともにmRNAの分解速度を速める効果が認められた。一方、乱流は転写には影響せずmRNAの安定化を起こす作用が確認された(Am J Physiol 2004)。内皮細胞に留まらず肝細胞においてもshear stressが遺伝子の発現を調節する、すなわちshear stressは転写因子Sp1とEts-1を介してPAI-1(プラスミノーゲン・アクチベータ阻害因子)の遺伝子の転写を活性化することが示された(Am J Physiol 2006)。最近開発したシリコンチューブ型流れ負荷装置による検討で内皮遺伝子の応答がshear stressとcyclic strainの同時負荷と各々の単独負荷では異なることを観察した(J Biotechnology 2008)。
クリックすると拡大します。Shear stressが内皮細胞の遺伝子の発現を転写調節あるいは転写後調節することを明らかにした。転写調節に関しては、VCAM-1遺伝子の発現がshear stressで抑制を受けるが、それには遺伝子プロモータに2個並んで存在する転写因子AP-1結合エレメント(TGACTCA)がshear stress応答配列として働いていることを示した(Am J Physiol 1997)。転写後調節に関しては、顆粒級・マクロファージ・コロニー刺激因子(GM-CSF)の遺伝子発現がshear stressで増加するが、その効果は転写ではなくmRNAの安定化を介していることを明らかにした(Cric Res 1998)。また既知の遺伝子だけでなく多くの未知の遺伝子もshear stressに感受性のあることをmRNAのdifferential display法で示した(BBRC 1996)。共同研究によりshear stressに反応するG蛋白受容体ファミリーに属する未知の遺伝子をクローニングした(BBRC 1997)。また、流れで誘発されるCa2+反応に関わるP2X4プリノセプターの発現がshear stressで抑制を受けるが、これは転写因子SP1が関連した転写抑制に基づくことを明らかにした(Am J Physiol 2001)。さらにDNAマイクロアレイ解析で遺伝子全体の約3%(約600の遺伝子に相当)がshear stressに応答することを観察した(J Athero Thromb 2003)。最近、遺伝子に対するshear stressの作用が層流と乱流で異なることを明らかにした。線溶と血管のリモデリングに関わるウロキナーゼ型のプラスミノーゲン・アクチベータ(uPA)の遺伝子の発現は層流で低下し、乱流で増加した。層流は転写因子GATA6を活性化し転写を抑制するとともにmRNAの分解速度を速める効果が認められた。一方、乱流は転写には影響せずmRNAの安定化を起こす作用が確認された(Am J Physiol 2004)。内皮細胞に留まらず肝細胞においてもshear stressが遺伝子の発現を調節する、すなわちshear stressは転写因子Sp1とEts-1を介してPAI-1(プラスミノーゲン・アクチベータ阻害因子)の遺伝子の転写を活性化することが示された(Am J Physiol 2006)。最近開発したシリコンチューブ型流れ負荷装置による検討で内皮遺伝子の応答がshear stressとcyclic strainの同時負荷と各々の単独負荷では異なることを観察した(J Biotechnology 2008)。
Shear stress の感知・情報伝達機構
内皮細胞がshear stressを感知して、その情報を細胞内部に伝達する機構に関して、セカンドメッセンジャーであるCa2+を介する情報伝達経路のあることを初めて明らかにした(In Vitro Cell Dev Biol 1988)。強い機械的刺激(バルーンによる摩擦)は単独で内皮細胞内にCa2+上昇反応を起こす(Biorheology 1994)が、弱い機械的刺激であるshear stressは細胞外ATPの存在を必要とし、とくにATP濃度が500 nM付近でshear stressの強さに依存したCa2+上昇反応の起こることを発見した(BBRC 1991)。このCa2+反応は流速依存性に増加する細胞膜へのATPの到達量の増加ではなく機械的刺激であるshear stressに依存することを流れ負荷に使う潅流液の粘性を変える独自の実験方法で確認した(BBRC 1993)。さらに、このCa2+反応が細胞の辺縁の局所から開始し、Ca2+波として細胞全体に伝搬して行くこと、この開始点はカベオリンが密に分布する場所であることから、流れ刺激の情報が細胞膜の陥入構造物であるカベオラから入力される可能性を示した(Proc Natl Acad Sci 1998)。また、共同研究で、こうしたCa2+を介する情報伝達にミオシン軽鎖キナーゼが深く関わっていることを明らかにした(FASEB J 1998)。肺動脈内皮細胞にATP作動性カチオンチャネルのP2X4が優勢的に発現し(Am J Physiol 2000)、それが流れ刺激で起こるCa2+ 流入に中心的な役割を果たすこと(Circ Res 2000)、さらにP2X4を介したCa2+ 反応に流れ刺激によって放出されるATPが関わることを示した(Am J Phsiol 2003)。P2X4遺伝子の欠損マウスを作製したところ、このマウスの内皮細胞ではshear stressによるCa2+ 流入反応が起こらずNO産生が抑制されていた。このため血流増加による血管拡張反応が障害され血圧が上昇していた。また、血流を変化させたときに生じる血管のリモデリングも障害を受けていた。このことから、P2X4を介する血流刺激の情報伝達は循環系の調節に個体レベルで重要な役割を果たしていることが示された(Nat Med 2006)。最近、shear stressによる内因性ATPの放出反応に細胞膜カベオラ・ラフトに存在するFoF1ATP合成酵素が関わっていることが判明した(Am J Physiol 2007)。
ムービーをご覧になるには、Quick Timeが必要です。下記のサイトよりダウンロードして下さい。
http://www.apple.com/jp/quicktime/download/