東京大学医学部生化学教室
東京大学大学院医学系研究科 分子細胞生物学専攻 生化学分子生物学 細胞情報部門
Department of Biochemistry and Molecular Biology, Faculty of Medicine, The University of Tokyo
研究紹介
研究の背景![]()
ベルリンでのシェリング賞授賞式講演内容(2000年9月27日)もご参照下さい
細胞情報学教室では、清水教授以下、中村准教授、石井准教授、北助教、進藤助教、高橋助手(共通機器担当)というスタッフ体制で、脂質メディエーター(lipid mediator)と呼ばれる種々の生理活性脂質の生体内における機能や、脂質メディエーターを作り出すもととなる、生体を構成する脂質膜の構築や再構築のメカニズムを明らかにするべく、様々なアプローチで研究を展開しています。
私たちが研究の対象としている脂質メディエーターの典型としては、エイコサノイド(eicosanoid)として知られる、プロスタグランジン類(prostaglandin, PG)やロイコトリエン類(leukotriene, LT)などがあります。これらはアラキドン酸(arachidonic acid)という不飽和脂肪酸に由来する一連の代謝産物であり、多くの場合、膜グリセロリン脂質にエステルとして存在するアラキドン酸に由来します。細胞外に放出されたエイコサノイドは、特異的な7回膜貫通型のGタンパク質共役型受容体(GPCR)に結合し、ホルモン様のシグナル分子として働くことが知られています。これらエイコサノイドの生理機能は多岐にわたり、炎症やガンなどに代表される病態への関与もよく知られております。_
一方、脂肪酸をひとつ失ったリン脂質、すなわち「リゾリン脂質」と呼ばれる脂質のうちあるものは、アセチル基の転移(付加)によって血小板活性化因子(platelet-activating factor, PAF)という脂質メディエーターを生じます。PAFもまた、特異的なPAF受容体(PAFR)を介して(血小板の活性化に限らない)多様な生理機能を発揮します。また、リゾホスファチジン酸やリゾホスファチジルコリン、スフィンゴシン-1-リン酸などのリゾリン脂質も脂質メディエーターとして働きます。
私たちの研究の大きな目標は、これらの脂質メディエーターがどのように作られ(脂質メディエーター産生酵素・脂質代謝物)、どのように標的に作用し(脂質メディエーター受容体/GPCR)、どのような生理機能を発揮するのか(細胞内シグナル解析・動物を用いた疾患モデル等)を明らかにすることであり、同時にこれら脂質メディエーター産生の背景となる、生体における脂質膜の構築・再構築のメカニズム(脂質代謝酵素)の一端を明らかにすることです。
研究課題 ![]()
- 脂質メディエーター受容体(GPCR)の研究(中村准教授・石井准教授)
- ホスホリパーゼA2の研究(北助教)
- PAF合成酵素の精製・クローニング(進藤助教)
- 脂質膜の構築・再構築のメカニズム(進藤助教)
- 質量分析による脂質メディエーターの同定・定量(北助教・高橋助手)
私たちの研究室で世界に先駆けてcDNAクローニングに成功したPAF受容体は、脂質をリガンドとするGPCRのcDNAクローニングとしても初めての例となりました(Honda, et al., Nature 349, 342-346, 1991; Nakamura, et al., J. Biol. Chem. 266, 20400-20405, 1991)。その後、横溝元助教授ら(2006年2月より九大医学部教授に着任)によりロイコトリエンB4受容体(BLT1)がcDNAクローニングされ、白血球の走化性に関わる受容体であることが明らかになりました(Yokomizo, et al., Nature 387, 620-624, 1997)。また、BLT1のプロモーター解析(ゲノム解析)の過程で明らかとなったロイコトリエンB4第2受容体(BLT2)の報告(Yokomizo, et al., J. Exp. Med. 192, 421-432, 2000)など、重要な脂質メディエーター受容体の同定に成功してきました。現在、当教室では、PAF受容体、ロイコトリエン受容体を中心に、それらの細胞における機能や、遺伝子改変マウス(KOマウス・Tgマウス)を用いた個体レベルでの機能解析を行っています。
石井助教授らによって樹立されたPAF受容体欠損マウス(Ishii, et al., J. Exp. Med. 187, 1779-1788, 1998)を用い、引地らはPAFと骨粗しょう症の関わりを示しました(Hikiji, et al., J. Clin. Invest. 114, 85-93, 2004)。同じく木原らはPAFが多発性硬化症(MS)の症状の増悪に関与している可能性を指摘するなど(Kihara, et al., J. Exp. Med. 202, 853-63, 2005)PAFの機能の多様性が明らかになってきています。
また、これまでリガンドの報告されていないGPCR(孤児受容体、orphan receptors)のうち、脂質をリガンドとすることが期待されるものについて、精力的なリガンド探索を行っています(デ・オーファニング・de-orphaning)。この最初の成功例は、野口らによるリゾホスファチジン酸(LPA)第4受容体、LPA4の発見です(Noguchi, et al., J. Biol. Chem. 278, 25600-25606,2003)。脂質をリガンドとする可能性の考えられたGPCRのうち、これまでの概念と全く異なる様式で活性化される2つの受容体G2A(Murakami, et al., J. Biol. Chem. 279, 42484-91, 2004)およびTDAG8(Ishii, et al., J. Biol. Chem., 280, 9083-87, 2005)を報告しました。これらの受容体は、低いpHによって活性化されるため「proton-sensing receptor」と考えられ、その生体における役割やシグナル伝達機構などを現在解析中です。また、奥野らによりBLT2の生体内リガンドが12-HHTであるという発見もしました(Okuno et al., J. Exp. Med 205, 759-766, 2008)。
当教室において魚住らにより細胞質型ホスホリパーゼA2α(cPLA2α)の欠損マウスが作製され(Uozumi, et al., Nature 390, 618-622, 1997)、炎症を伴う様々な病態モデルにおいてエイコサノイドやPAFの産生を介して病態の増悪に関わることを示してきました。ほ乳類においてホスホリパーゼA2は20種以上知られていますが、アラキドン酸カスケードにおいては、cPLA2αが中心的役割を担っていることが明らかとなりました。
cPLA2β、cPLA2γというパラログがその後ESTデータベースから同定されたことを踏まえ、大戸らはこれらの既知アミノ酸配列の部分的保存性をもとにゲノムデータベースから新たにcPLA2δ、cPLA2ε、cPLA2ζを推定、同定し報告しました(Ohto, et al., J. Biol. Chem., 280, 24576-83, 2005)。既知cPLA2分子に加え、これらの新規cPLA2の酵素学的性質と生体内での機能を解析しています。cPLA2α欠損マウスは多くの病態モデルで、その症状が軽微であることが明らかとなりました(Shimizu, et al., IUBMB Life, 58, 328-333, 2006; Shimizu, Annual Rev. Pharmacol. Toxicol 49, in press, 2009)
リゾPAFにアセチル基を転移してPAFを合成するリゾPAF アセチル転移酵素は長年その酵素学的特徴についての研究や、酵素タンパク質の精製の試みがなされてきましたが、この酵素は膜タンパク質であるため精製が非常に困難であり、これまでその精製およびcDNAクローニングの報告はなされておりません。進藤助教らによりこの酵素の研究が進められています(Shindou, et al., J. Immunol., 175, 1177-1183, 2005)。2007年にこの研究で大きな飛躍がありました。リゾPAFアセチル転移酵素のクローニングに成功しました(Shindou, et al. J. Biol. Chem, 282, 6532-6539, 2007; Harayama, et al., J. Biol. Chem, 283, 11097-11106, 2008)。クローニングした酵素はかつてマクロファージで観察されたような種々の性質(カルシウム要求性、エンドトキシンによる活性化と誘導、抗炎症ステロイドによる誘導の阻害などの性質を持っていました。
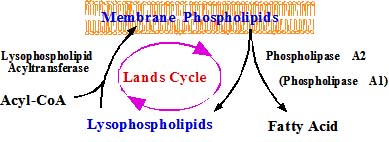
上述のリゾPAFアセチル転移酵素の研究は、膜グリセロリン脂質の構築・再構築に関わる酵素に関する研究と深く関わっています。細胞膜リン脂質のsn-1・sn-2脂肪酸の非対称性は古くから知られており、これにはリン脂質リモデリング酵素が関わっていると考えられてきましたが、その実態はまだまだ明らかではありません。当教室では進藤助手らによりリン脂質のリモデリングに関わるような酵素を同定するべくその探索が行われています。最近、リゾホスファチジルコリンにアシル基を転移する二種類のアシル転移酵素のクローニングに成功しました(Nakanishi et al., J. Biol. Chem., 281, 20140-7, 2006)。前者は肺胞2型上皮細胞に高発現しており、肺サーファクタント産生に関係する可能性があり、もう一つの酵素は炎症細胞に発現し、リン脂質膜を合成すると共に、PAFの産生に関わることが明らかとなりました(Shindou, et al. J. Biol. Chem, 282, 6532-6539, 2007)。sn-2位の脂肪酸が代謝回転する現象を見出したのは、William E. Lands(1960年)でランズ回路と呼ばれています(下図)。40年経過して、その酵素を発見することが出来たと思っています。アミノ酸活性化酵素は20種類あることが知られていますが、リゾリン脂質アシル転移酵素は何種類存在し、脂肪酸とグリセロリン脂質の極性基を認識し、どの様に秩序立った生体膜を形成するかが大きな研究課題となりました (Hishikawa et al., PNAS, 105, 2830-2835, 2008)。
私たちが研究対象とする生理活性脂質を同定・定量するためには、高感度で信頼性の高い分析手法が不可欠です。HPLCとエレクトロスプレーイオン化質量分析計を組み合わせたいわゆるLC/MSの手法の開発を行っています。リン脂質や過酸化脂質などの解析については、メタボローム寄付講座(田口教授)との連携を行っています。エイコサノイドおよびPAFの一斉定量系は北・高橋らによって構築され(Kita, et al., Anal. Biochem. 342, 134-43, 2005)、各種疾患モデル動物実験などに活用することにより、包括的に生体内でのエイコサノイド産生の意義を検討するための材料を提供しています(Yoshikawa, et al., J. Biol. Chem. 281, 14663-9, 2006)。
脂質メディエーターの研究から、膜の脂質合成がどの様に調節され、再構築されるかという大きな生物学的問題へと研究は発展しています。
教室の研究手技 ![]()
我々の教室は生化学・分子生物学講座であり、基本的には生化学・分子生物学的手法での研究を進めていますが、同時に細胞の生理機能(カルシウム応答、遊走、形態変化など)を観察し、また、発生工学的手段で分子の機能を個体で観察することを行っています。教室で行われている手技はラボマニュアルの通りですが、以下のようにまとめることが出来ます。
- 生化学
- 遺伝子工学
- 細胞生物学と細胞生理
タンパク精製、抗体作製、ペプチドシークエンシング、酵素精製、酵素活性測定、ウエスタンブロッティング、脂質の抽出と精製(HPLC、TLC、ガスクロマトグラフィーなど)、質量分析計(Thermo Electron TSQ 7000, Thermo Electron TSQ Quantum Ultra, Shimadzu Axima DFR, Applied Biosystems 4000 Q TRAP, Waters Q-Tof microなど)、Biacore 3000による分子間相互作用の解析
クローニング、各種ハイブリダイゼーション、自動塩基決定装置、定量的PCR、yeast two-hybrid screening、in situ hybridization等
各種クローン化細胞の培養、ニューロン、グリアなどの初代培養。細胞内セカンドメッセンジャーや各種リン酸化酵素の測定。細胞内カルシウム濃度測定(CAF-110, Argus CA-50、FLEX Station)。微分干渉顕微鏡、蛍光顕微鏡、共焦点顕微鏡。マウス樹状細胞、T細胞の機能解析。
熱意あふれた大学院生募集・ポストドク募集 ![]()
細胞情報学講座では上記のテーマに興味を持ち、「熱意、体力と協調性のある大学院生」を募集しています。毎年、8月上旬に願書締め切りがあり、10月中旬に入学試験(英語、専門科目)があります。質問は清水までどうぞ。なお、応募資格は医学部、歯学部、獣医学部卒あるいは4年生他学部の修士課程修了(見込み)の方です。また、平成11年より医科学修士学生(合計26名)を募集しています。この試験は8月下旬です。応募資格は普通4年制の大学卒業者(文系、理系を問わず)で、6年間一貫教育することを目標にしています。詳細は医学部大学院係にお問い合わせ下さい。
ポストドク募集の連絡は改めて行います。
- 教育方針
- リサーチセミナー:研究報告セミナー
- 文献セミナー
- ジャーナルクラブ (Journal Club, JC)
- 輪読会
- ゲストセミナー
- 研究テーマ
基本的には一人一つのテーマで、生化学、分子生物学から細胞生物学まで幅広い技術と考え方を身につけるよう指導しています。最初からキットの世界に入らないよう、生化学的な教育を重視したいと思っています。この教室の一つの特徴は生化学教室でありながら、技術的に非常に幅広い蓄積があることです。実技はスタッフがマンツーマンで指導します。その他に週に3回セミナーがあり、幅広い知識と考え方を身につける練習をします。
毎週火曜日午前9時-12時(一人1時間、3名);配付資料は英語です。討論の座長は院生が行います。
毎週火曜日12時-13時(研究セミナー終了後、昼食を食べながら)数個の論文を批判的に読むのが目的です。
メール配信にて主要雑誌の内容紹介を交代でしています。1回に4誌を報告します。担当はD3の安田です。
毎週土曜日の正午-1時まで、代表的教科書の輪読会をしています。現在は未定 です。問い合わせは真砂まで。
平均して月に一回程度、ゲストセミナーを行います。外国人が講師の場合は、大学院生が司会を行います。
文献セミナー、JC、輪読会は誰でも自由に参加できます。現に他教室や他学部学生等が出席しています。希望者は清水まで連絡を下さい。
生化学・分子生物学の一定の修練の後、神経、生体防御、免疫、細胞内シグナル伝達、リン脂質の代謝学、受容体生化学など、興味に応じて各自のテーマを決定します。大学院の4年間、あるいは6年間で基礎的な思考や技術を身につけ、その後の研究生活の土台を作るのが目的です。
参考論文リスト ![]()
- PAF受容体のクローニング (Honda, et al., Nature 349, 342-346, 1991)
- ヒトPAF受容体のクローニング (Nakamura, et al., J. Biol. Chem. 266, 20400-5, 1991)
- ニューロンにおけるPAF受容体の機能 (Bito, et al., Neuron 9, 285-294, 1992)
- PAF受容体過剰発現マウスの表現型解析 (Ishii, et al., Embo J. 16, 133-142, 1997)
- ロイコトリエンB4第一受容体(BLT1)のクローニング (Yokomizo, et al., Nature 387, 620-624, 1997)
- cPLA2欠損マウスの表現型解析 (Uozumi, et al., Nature 390, 618-622, 1997)
- PAF受容体欠損マウスの表現型解析 (Ishii, et al., J. Exp. Med. 187, 1779-1788, 1998)
- BLT1の転写機構の解明 (Kato, et al., J. Exp. Med. 192, 413-420, 2000)
- ロイコトリエンB4第二受容体(BLT2)のクローニング (Yokomizo, et al., J. Exp. Med. 192, 421-432, 2000)
- cPLA2欠損マウスのARDSモデルにおける解析 (Nagase, et al., Nat. Immunol. 1, 42-46, 2000)
- cPLA2欠損マウスの肺線維症モデルにおける解析 (Nagase, et al., Nat. Med. 8, 480-484, 2001)
- cPLA2欠損マウスの血小板機能における解析 (Wong, D. et al., J. Exp. Med. 196,349-57,2002)
- オーファン受容体リガンド探索プロジェクトの最初の成功例(新しいLPA受容体の発見)(Noguchi, et al., J. Biol. Chem. 278, 25600-6,2003)
- cPLA2の骨代謝、関節リウマチへの関与 (Hegen, et al., J. Exp. Med. 197, 1297-1302, 2003; Miyaura, C. et al., J. Exp. Med. 197, 1303-10, 2003)
- PAF受容体と閉経後骨粗鬆症の関係 (Hikiji, et al., J. Clin. Invest. 114, 85-93, 2004)
- G2Aがプロトン感受性GPCRであることを発見 (Murakami, et al., J. Biol. Chem. 279, 42484-91, 2004)
- TDAG8がプロトン感受性GPCRであることを発見 (Ishii, et al., J. Biol. Chem. 280, 9083-7, 2005)
- エイコサノイド一斉定量系の開発 (Kita, et al., Anal. Biochem. 342, 134-43, 2005)
- 3種の新規cPLA2の同定 (Ohto, et al., J. Biol. Chem. 280, 24576-83, 2005)
- LPSによるPAF合成酵素の活性化の解析 (Shindou, et al., J. Immunol. 175, 1177-83, 2005)
- PAFが多発性硬化症に関連 (Kihara, et al., J. Exp. Med. 202, 853-63, 2005)
- ラット海馬における脂質メディエーター産生 (Yoshikawa, et al., J. Biol. Chem. 281, 14663-9, 2006)
- 肺サーファクタント脂質合成酵素の単離 (Nakanishi, et al., J. Biol. Chem. 29, 20140-7, 2006)
- cPLA2の生化学的特徴及び生理学的機能 (Shimizu, et al., IUBMB Life 58, 328-33, 2006)
- アセチルCoA:リゾPAFアセチル転移酵素のクローニング (Shindou, et al., J. Biol. Chem. 282, 6532-39, 2007)
- 生体膜の非対称性と多様性に寄与するリゾリン脂質アシル転移酵素群の同定 (Hishikawa, et al., PNAS 105, 2830-5, 2008)
- 非炎症時に働くPAF合成経路の同定 (Harayama, et al., J. Biol. Chem. 283, 11097-106, 2008)
- 12-HHTがBLT2の生体内リガンドであることを発見 (Okuno, et al., J. Exp. Med. 205, 759-66, 2008))
- ヒト病態における生理活性脂質 (Shimizu, et al., Annual Rev. Pharmacol. Toxicol. 49, 2009))