B. これまでの成果と今後の展望
1. 背景
(ア) AMPA受容体(グルタミン酸受容体サブタイプ)
- ①グルタミン酸受容体はイオンチャネル型と代謝調節型がある。AMPA受容体はNMDA受容体、カイニン酸受容体などと共に前者に属す。
- ②AMPA受容体は速い神経伝達を制御する神経細胞の興奮性を規定するグルタミン酸受容体で、中枢神経に広く分布している。
- ③AMPA受容体は4種類のサブユニット(GluA1~ GluA4)の様々な組み合わせに依る四量体構造をとり、Na+/ K+を透過させることで、膜電位を制御している。
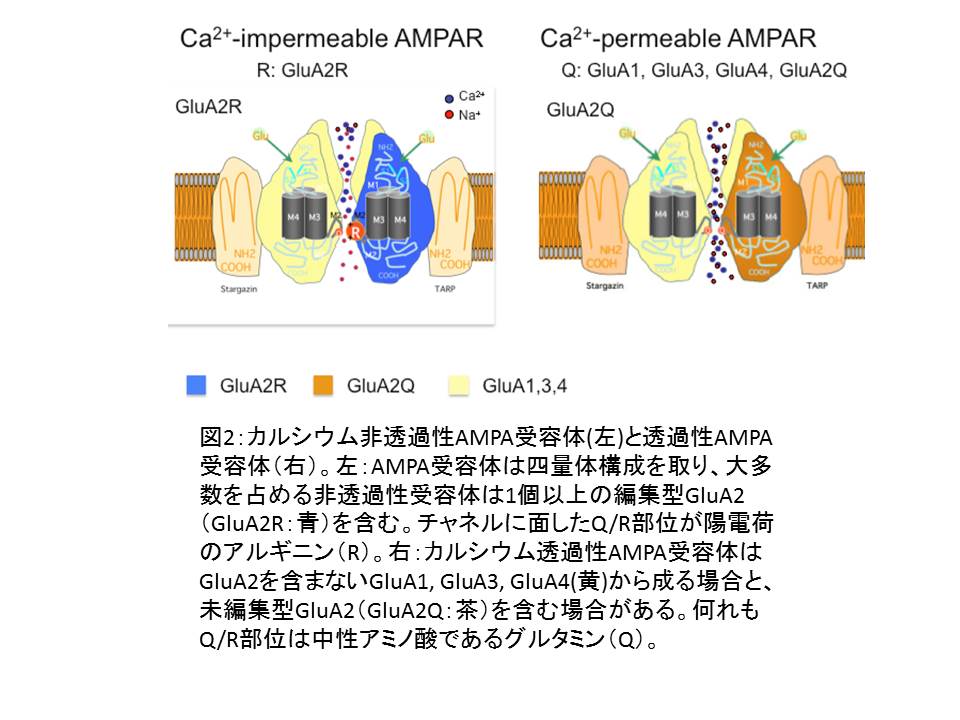 図2
図2
- ④サブユニット構成によりCa2+ 透過性のものと非透過性のものがある(図2)。Ca2+非透過性AMPA受容体が圧倒的大多数を占める。Ca2+ 透過性AMPA受容体の過剰発現は動物にてんかんを引き起こすことが知られていた。
(イ) GluA2サブユニットとそのRNA編集(RNA editing) (図2)
- ①AMPA受容体とCa2+ 透過性
- 1. AMPA受容体のCa2+透過性はGluA2がサブユニットに含まれるかどうかで決まる。正常のニューロンが発現するGluA2は全て編集型(②で後述)である。
- 2. Ca2+非透過性AMPA受容体:ニューロンに発現するAMPA受容体の大多数を占め、編集型GluA2 をサブユニットに含む
- 3. Ca2+透過性AMPA受容体:サブユニット構成にGluA2を欠くもの(生理的には海馬の抑制性ニューロンやグリア細胞)と、未編集型GluA2を含むもの(生理的にはグリア細胞の一部に存在するとされる)とがある。
- ②未編集型GluA2の発現の意味するもの
- 1. GluA2はグルタミン・アルギニン部位(Q/R部位)でRNA編集5を受ける。
- 2. すなわち、GluA2 pre-mRNAエクソン11にあるグルタミン(Q)コドン(CAG)のアデノシンがイノシンに置換され、翻訳時にイノシンはグアノシンとして認識されるので、アルギニン(R)(CGG=CIG)に翻訳される。ゲノム情報CAGがタンパクレベルではR(CGG)に置換される。
- 3. Q/R部位はチャネルに面しており、陽荷電のRはCa2+ の透過をブロックするが、中性のQはブロック出来ない。
- 4. AMPA受容体の他の3種類のサブタイプのQ/R部位にはRNA編集が起こらず、Q型のため、GluA2をサブユニットに持たないAMPA受容体やGluA2がサブユニットに含まれていても未編集の場合、AMPA受容体のCa2+ 透過性は高い。
- 5. ヒトを含む哺乳類ではGluA2のQ/R部位は100%編集され、GluA2の発現もAMPA受容体サブユニットの過半を占めるため、ニューロンに発現するAMPA受容体は殆どがCa2+非透過性である。
- 6. GluA2Q/R 部位にRNA編集が起こらないことで
- a) AMPA受容体のチャネル特性が変わる(Ca2+透過性になる)。
- b) 未編集型GluA2は編集型GluA2に優先してシナプス後膜へ輸送される。
- c) そのため、未編集型GluA2が発現した分、機能的AMPA受容体に占めるCa2+透過性AMPA受容体の割合が増す。(ニューロンへのCa2+流入は未編集型GluA2が100%になると野生型(全て編集型)の29倍、50%でも4〜5倍に上昇するとされている6。)
- d) GluA2のRNA編集が出来ない変異動物はてんかん重積のため、生後3週以内に死亡してしまう7, 8。
- 7. したがって、未編集型GluA2の発現自体が少量であってもニューロン毒性をもつ。
(ウ)RNA編集酵素ADAR2 (adenosine deaminase acting on RNA 2)
 図3
図3
- ① RNA編集部位は2200万カ所以上にあるとされている。哺乳類の中枢神経ではRNA編集が極めて盛んに行われており、その大多数はアデノシン・イノシン(A-I)置換である。
- ② このA-I置換は二重鎖RNAに働く酵素により行われ、哺乳類では3種類のアイソフォーム(ADAR1-ADAR3)が知られている。
- ③ GluA2 Q/R部位のA-I置換は特異的にADAR2により触媒され、ADAR2の欠損ではこの部位のRNA編集は起こらない。
- ④ ヒトでの検討でADAR1,ADAR2とも中枢神経では主にニューロンに発現し、A-I置換に関與しているが、ADAR3は中枢神経特異的な発現パターンを持ちながら、発現は主としてグリアで、A-I置換活性も明らかでない9。