> Home > 研究室での取り組み

オートファジー(自食作用)
生命を維持するためには、一旦合成したタンパク質を適切に分解処理する必要があります。ユビキチン・プロテアソーム系が細胞質における選択的タンパク質分解の主役であるのに対し、オートファジーは細胞質成分をリソソームで分解するための主要分解機構です。オートファジーはタンパク質だけではなくミトコンドリアなどのオルガネラをも分解できる大規模なシステムで、酵母から高等動植物に至るまで非常によく保存されています。オートファジーでは約1μmの領域がランダムに包み込まれるため、オートファジーは原則として非選択的な分解システムであると考えられてきましたが、最近になって一部の基質は選択的に取り込まれることもわかってきています。
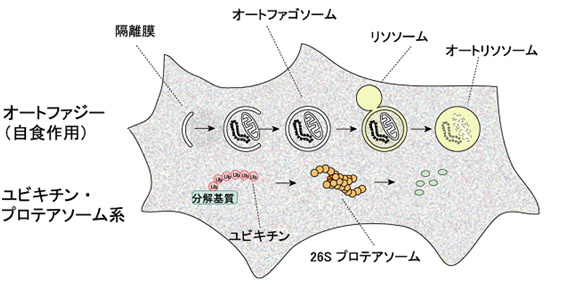
図1 真核生物の主要な細胞内分解機構
(上段)オートファジーではまず細胞質の一部が隔離膜によって取り囲まれてオートファゴソームが形成される。次にオートファゴソームの外膜がリソソーム膜と融合しオートリソソームとなり、内容物が分解される。
(下段)ユビキチン・プロテアソーム系では、ユビキチン鎖で標識された基質が、プロテアソームによって分解される。これは厳密な認識機構をもった選択的タンパク質分解系である。
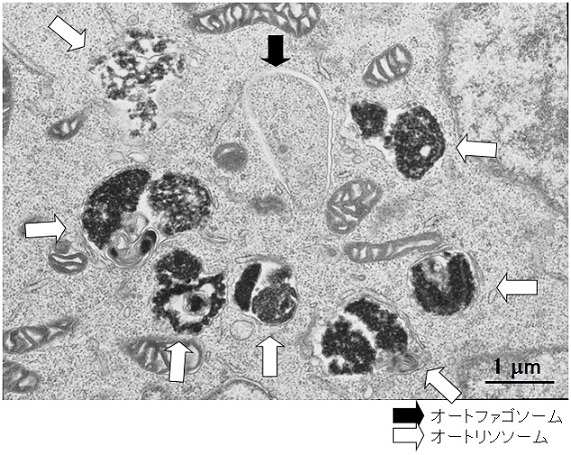
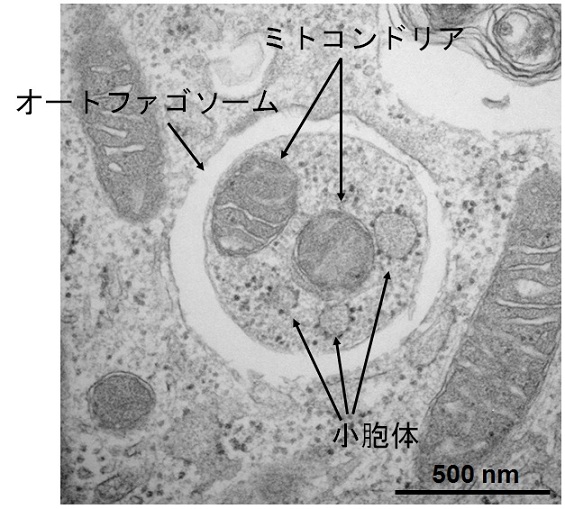
図2 オートファジーの電子顕微鏡像
(上段)栄養飢餓状態のマウス胚性幹細胞(長浜バイオ大学 山本章嗣博士撮影)
(下段)栄養飢餓状態のマウス線維芽細胞(東京医科歯科大学 岸千絵子博士撮影)
これまでの研究成果
1 オートファジーの分子機構
オートファジーの分子機構は大隅良典教授(現・東工大)のグループを中心にした出芽酵母の研究によって大きく進展してきました。特にオートファゴソーム形成に関わるAtg分子群の解析がこの分野の突破口となりました。これらの中には、ユビキチン化と類似した特殊な翻訳後修飾システムがふたつも存在していたなど、驚くべき仕組みが隠されていました(補足図1)。
私たちはこれらの酵母Atgタンパク質のホモログを足がかりに、哺乳類細胞でのオートファジーの分子機構について解析してきました。特に、栄養シグナル伝達因子、オートファゴソーム形成因子、オートファゴソーム・リソソーム融合因子の分子細胞生物学的解析を行っています。FIP200、Atg101、シンタキシン17などの多細胞生物独自の分子装置も発見されています。
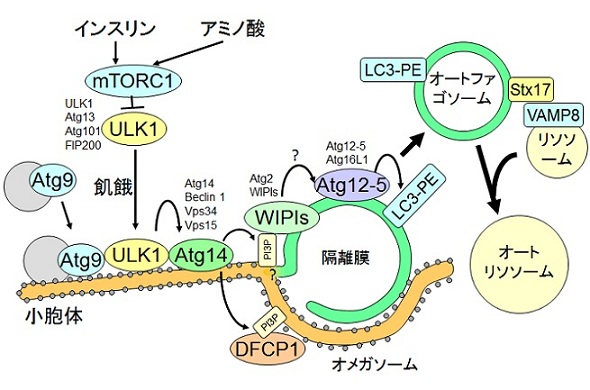
図3 哺乳類細胞でのオートファジー分子機構のモデル図
アミノ酸やインスリンはmTORC1複合体を介してオートファジーを抑制している。mTORC1はULK1-Atg13-FIP200-Atg101複合体と直接結合してオートファジーを抑制する。飢餓時にはULK1複合体は小胞体膜(あるいはその近傍)へAtg9小胞とともに移動 し、その下流ではAtg14-Beclin 1-Vps34-p150複合体、Atg2-WIPI複合体、Atg12-Atg5-Atg16L1複合体、LC3 (Atg8ホモログ)などがオートファゴソーム形成において重要な役割を担う。その後完成したオートファゴソームにシンタキシン17がリクルートされ、リソソームとの融合を誘導する。
GFP-Atg5のムービー(1)(60倍速)4.2 MB Quick Time
GFP-Atg5のムービー(2)(60倍速)600 KB avi
2 哺乳動物個体でのオートファジーモニター法の開発
これまでオートファジーの検出は電子顕微鏡観察に大きく依存していました。そこで、より簡便かつ特異的にオートファゴソームを検出する目的で、全身のオートファゴソームが蛍光標識されるトランスジェニックマウス(GFP-LC3マウス)を作製しました。このマウスを用いることで、オートファジーは栄養飢餓に応じて神経系を除くほぼすべての臓器で顕著に活性化されることが確認されました。さらに、胸腺上皮細胞などの一部の臓器では恒常的に高いレベルのオートファジーをおこしていることも観察されました。発生過程では、受精直後にオートファジーが活発化することもわかりました。これらのことからオートファジーは単に日常的な代謝回転のみならず、飢餓応答、発生、さらには各組織に特化した特殊な役割を果たしている可能性が示唆されました。
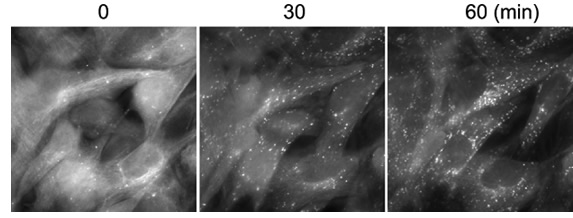
図4 培養細胞を用いた可視化の例
オートファゴソームを標識するGFP-LC3を発現するマウス線維芽細胞をアミノ酸飢餓培地に移してから観察した例。小さなドットがオートファゴソームを示す。

図5 GFP-LC3トランスジェニックマウスを用いた観察例
骨格筋では飽食時オートファジーはほとんどおこっていないが、24時間絶食後に著明に誘導される。肝などの他の臓器でも同様に誘導される。(このマウスは理研バイオリソースセンターから入手可能。下記参照)
3 哺乳動物でのオートファジーの生理的意義の解明
哺乳類におけるオートファジーの生理的役割を明らかにするために、オートファジー能を欠損するさまざまなマウスを作製しています。これまでの解析から、オートファジーは、新生児期に代表される飢餓適応、着床前の初期胚発生、細胞内浄化、神経変性抑止、腫瘍発生抑制に重要であることがわかっています。さらに、共同研究によって、オートファジーは細胞内侵入細菌の除去や抗原提示などのさまざまな機能があることもわかってきています。
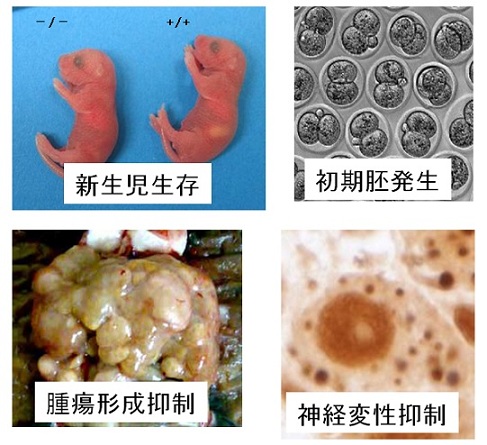
図6 オートファジーのさまざまな生理的役割の例
A.オートファジーは飢餓適応に重要である(Atg5欠損マウス(全身型)を用いた解析)
飢餓で誘導されるオートファジーの意義についてはオートファジーの能力を全身で欠損したマウス(Atg5ノックアウトマウス)新生児の解析から一部明らかになっています。このマウスはほぼ正常に生まれるものの、生後まもなく深刻なアミノ酸不足に陥ります(解説へ)。マウス胎児は胎盤から栄養供給をうけていますが、出生とともに突然飢餓にさらされます。正常マウスは全身でオートファジーを活性化することで自己タンパク質からアミノ酸を産生し、この飢餓に適応していると考えられます。
B.オートファジーは初期胚発生に必要である(卵特異的Atg5ノックアウトマウスの解析)
オートファジーは受精4時間後頃より活性化されます。この時期にオートファジーをおこせないマウス胚は、4-8細胞期で発生が停止してしまいます(解説へ)。これらの胚では新規タンパク質合成が低下しているため、マウス胚は母性タンパク質をオートファジーで分解することによって、この時期に必要なアミノ酸を獲得していると考えられます。
C.オートファジーには細胞内浄化作用・神経変性抑制作用がある(神経系特異的Atg5ノックアウトマウスの解析)
オートファジーには細胞内のクリアランス機能もあります。これは誘導的オートファジーではなく、定常的オートファジーの機能であると考えられます。これがうまく機能しないと、細胞内に異常なタンパク質が蓄積してきます。例えば、神経特異的Atg5ノックアウトマウスは神経変性疾患を発症します(解説へ)。これはさまざまな神経変性疾患や老化とも関連する重要な機能です。実際、ヒトの神経変性疾患のひとつ(SENDA病)では、WIPI4遺伝子の変異がみつかっています(解説へ)。
D.オートファジーには腫瘍発生抑制作用がある(Atg5モザイク欠損マウスの解析)
ATG5遺伝子を全身臓器でモザイク状に欠損するマウスを作成し、オートファジー欠損細胞の運命を全組織で長期間追跡しました。その結果、このマウスは肝でのみ多発性腫瘍を発生することが明らかとなりました(解説へ)。腫瘍細胞では、腫脹した不良ミトコンドリアの蓄積、異常な酸化ストレス応答、ゲノム傷害応答が観察された。これはオートファジーによる不良ミトコンドリアや毒性タンパク質の除去不全の結果であると予想されます。
当研究室の課題
1.オートファジーの生理的意義
オートファジーは一部のタンパク質やミトコンドリアなどのオルガネラを特異的に識別して分解することができることが最近の研究から判っており、このような選択的オートファジーの役割は基質の性質を考えることで理解できます。しかし、ほとんどの基質は実際はランダムに分解されており、この「非選択性」には、種を超えて保存されているオートファジーの基本的な秘密があると考えています。これらを総合して、分野横断的研究を展開し、オートファジーの真の役割の理解に迫ります。
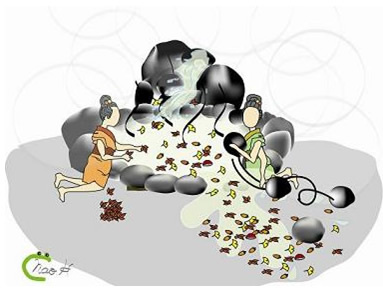
図7 オートファジーとユビキチン・プロテアソーム系の温泉モデル
ユビキチン・プロテアソーム系はゴミを個別に認識する(左)。一方、オートファジーは基本的には個別な識別をせずに、ゴミとともに湯も捨て、新しい湯を入れることで新鮮さを保つ(右)。一見無駄に思える方法でも、効率はよいと考えられる(画・細川奈生(OG))
2.制御メカニズム
オートファジーの特徴のひとつはそのダイナミックな調節にあります。細胞内ではmTORがオートファジーの主要な制御因子であることがわかっていますが、生体内ではどのような情報がオートファジーを本当にコントロールしているかはまだ明らかになっていません。さらには細胞がアミノ酸などの栄養素を感知する方法についてもわからないことがたくさんあります。オートファジーを研究対象の中心にしながら、生体や細胞の栄養シグナルを明らかにすることを目指しています。
3.オートファゴソーム形成の分子基盤
オートファジーの不思議な膜動態はまだまだ解明されていません。私たちは、特に、(1)オートファゴソーム形成部位の解明、(2)選択的基質のリクルート機構の解明、(3)隔離膜の伸長メカニズムの解明、(4)オートファゴソームとリソソームの融合機構の解明、を目指しています。哺乳類細胞には独自の分子機構があることもわかってきましたので、哺乳類細胞を用いてこれらの問題を解いていきたいと考えています。
4.モニター系の開発
GFP-LC3マウスの開発は有効な研究ツールを提供しましたが、まだ完全ではなく、いろいろな問題点も含んでいます。より普遍的なオートファジーの診断、定量方法の開発を試みています。
5.疾患とのかかわり
オートファジーを検出できるマウス、オートファジー機能を抑制したマウスなど、さまざまな研究ツールが揃いました。これらを用いて、オートファジーが関わる疾患、あるいはオートファジーを用いて制御することのできる疾患などを探しながら研究していきたいと思います。最近、ヒトの神経変性疾患のひとつ(SENDA病)でWIPI4遺伝子の変異がみつかりました(解説へ)。
当研究室の研究ツールは広く分与していますので、ご入り用の方は下記のリンクよりお申し込みください。
Riken BioResource Center Cell Bank
Riken BioResource Center Experimental Animal Division
Addgene
さらに詳しく知りたい方へ
オートファジーの生理機能・病態との関連
Mizushima, N., Komatsu, M.
Autophagy: renovation of cells and tissues.
Cell 147:728-41 (2011).
オートファジーの分子機構
Mizushima, N., Yoshimori, T., Ohsumi, Y.
The Role of Atg Proteins in Autophagosome Formation.
Annu. Rev. Cell Dev. Biol. 27:107-32 (2011).
オートファジーの検出方法
Mizushima, N., Yoshimori, T. and Levine, B.
Methods in mammalian autophagy research.
Cell 140; 313-326 (2010)
一般書
(水島昇著PHPサイエンス・ワールド新書 2011年)