ロイコトリエンB4(LTB4)はアラキドン酸に由来する生理活性脂質で、IL-8と並んで白血球を強く活性化する因子として知られています。nMという極めて低い濃度で細胞膜表面に存在する受容体に結合し、白血球の遊走、活性酸素の産生、ライソゾーム酵素の放出を促し、炎症反応や細菌感染に対する防御、といった生体にとって重要な役割を演じていると考えられてきました。また、乾癬や炎症性腸疾患、喘息といった疾患の発症因子であるとも考えられ、LTB4受容体拮抗薬は新しい抗炎症薬や喘息治療薬として開発が進められています。
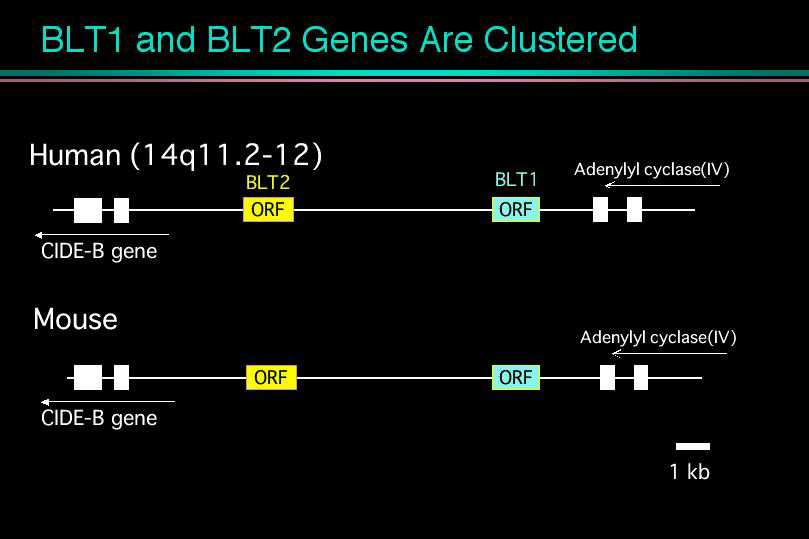
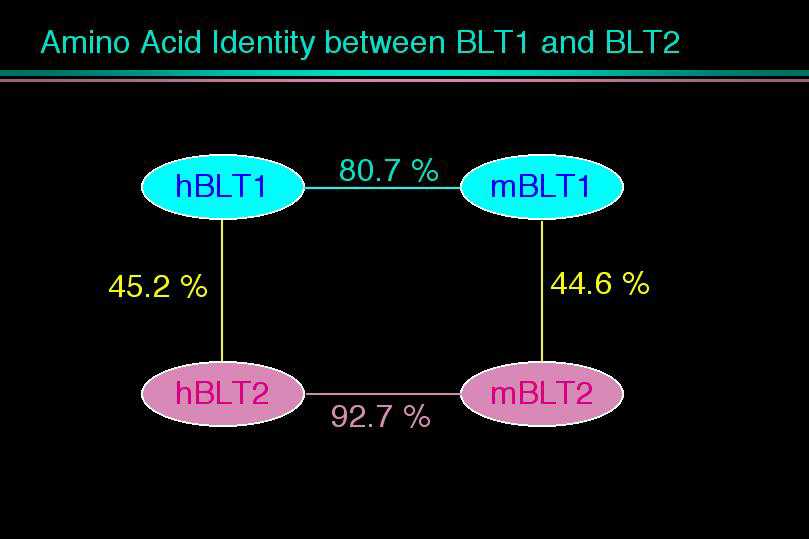
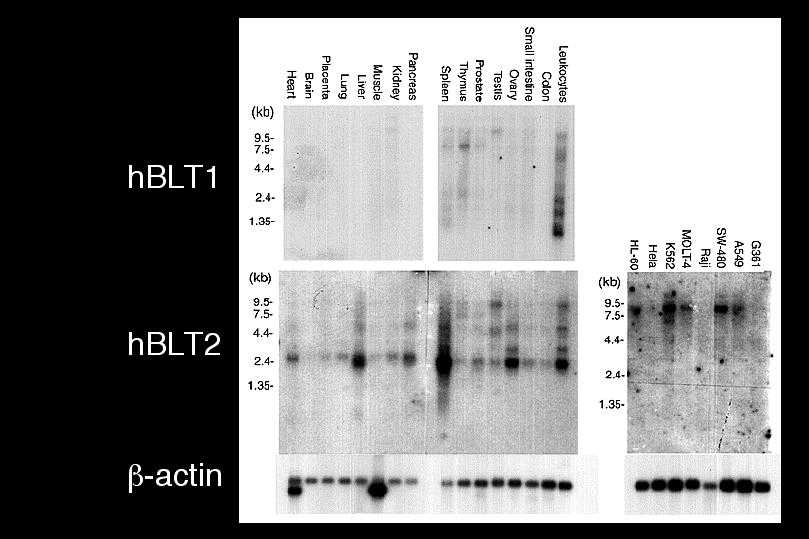
我々は1997年に世界で最初のロイコトリエン受容体の発見となったBLT1のcDNAクローニングをNature誌に発表しました。BLT1は白血球に強い発現が見られた反面、その他の臓器には発現しておらず、極めて厳密な臓器特異的な転写調節を受けていることがわかりました。そのメカニズムを解明する目的で、ヒトやマウスからBLT1のゲノムをクローン化し、プロモーター部位の解析を行いました。ヒトBLT1遺伝子はヒト14番染色体に存在していました。遺伝子の配列を読んでいくうちに、我々はおもしろいことに気づきました。BLT1遺伝子の上流にBLT1とよく似た受容体をコードするタンパク質翻訳領域が存在したのです。マウスの遺伝子でも、全く同じ位置に、ほとんど同じ構造を持った受容体がありました。BLT1はこれまで単離された他の受容体とはかなり違った一次構造を取っており、最も相同性の高かったソマトスタチン受容体と28%のアミノ酸同一性をもつに過ぎませんでした。しかし今回見つかった受容体はBLT1と45%という高い相同性を有しており、新たなエイコサノイドの受容体ではないかとの期待を抱かせました。
このように、遺伝子から見つかってきた受容体はOrphan receptor(みなし子受容体)と呼ばれます。リガンドが見つかって初めてその価値が見いだされるので、我々も早速この受容体のリガンドを探すことにしました。といっても、事は簡単ではありません。特異的な抗体もありませんから、受容体のアミノ末端に抗体が認識できるエピトープ(今回はHA-tag)を付けた発現ベクターを作成し、CHO細胞に遺伝子導入し、抗HA抗体でスクリーニングして安定発現株を3つとりました。これらの細胞に様々なリガンドをかけて細胞内のシグナルが動かないかどうかを観察しました。
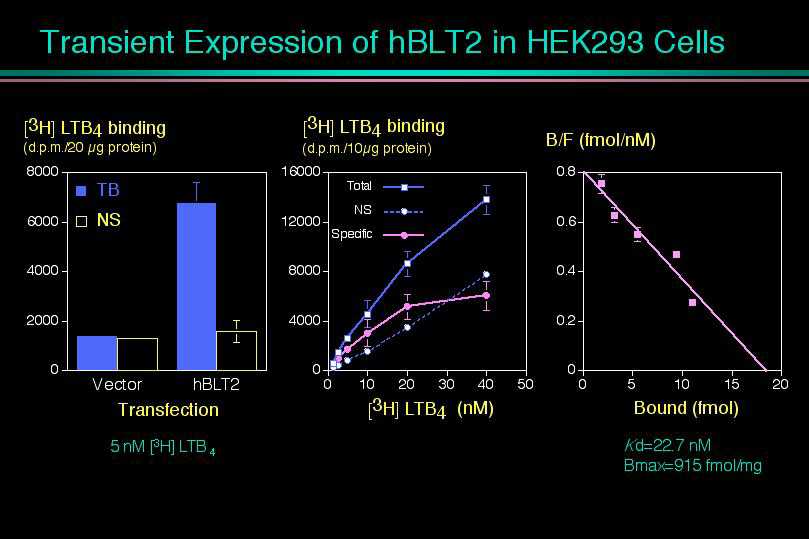
30種類くらいのリガンドを試したところ、何と驚くべき事に、この受容体もまたLTB4を認識して、細胞内カルシウム上昇、アデニル酸シクラーゼの阻害、細胞走化性を示しました。あわててHEK293細胞に一過性に発現して、細胞膜画分を使って結合実験を行うと、やはり、LTB4に対してKd値22 nMの特異的結合を示し、LTB4の低親和性の受容体であることがわかりました。そこで我々はこの受容体をBLT2と名付けました。ノザンブロッティングで発現臓器を調べると、BLT1とは大きく異なっており、脾臓に最大の発現がみられ、肝臓や卵巣をはじめ、ほとんど全ての臓器に広範囲な発現が観察されました。脾臓は、過去にマウスやヒトで低親和性のLTB4結合タンパク質の存在が報告されている臓器であり、今回クローニングしたBLT2が、過去に様々な議論のあった低親和性LTB4受容体の本体であると考えられます。すぐ近傍に二つの受容体、しかも、同じリガンドを認識する受容体が存在するという事実は、これらの受容体が進化の過程で「Gene duplication」を生じ、そのまま保存されてきたことを意味します。さらに今回見つかったBLT2のヒトとマウスの相同性が極めて高いと言う事実は、BLT2こそ進化の過程で保存される必要性が高かったことを示唆します。
さらに興味深いことが見つかりました。
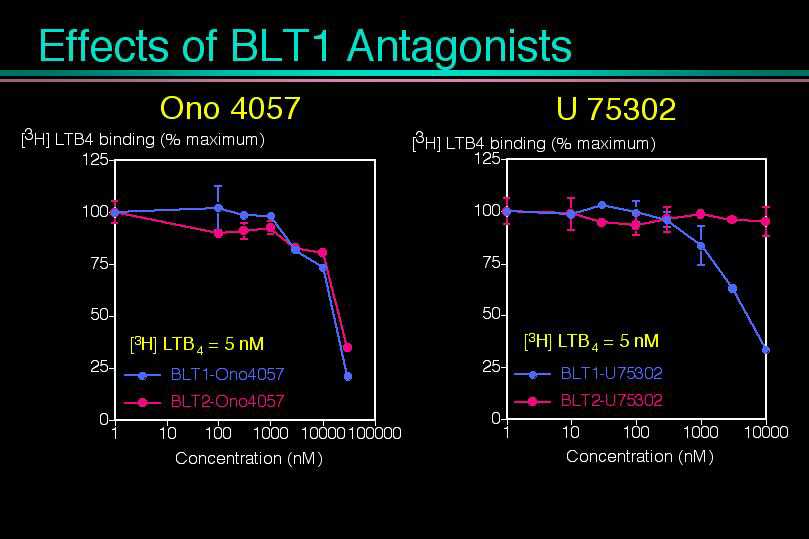
1)BLT2へのLTB4の結合は、これまで開発されてきたBLT拮抗薬の一部では阻害されないこと。これによってBLT2は薬理学的にBLT1とは異なった受容体であることがわかり、新しい創薬のターゲットになり得ます。
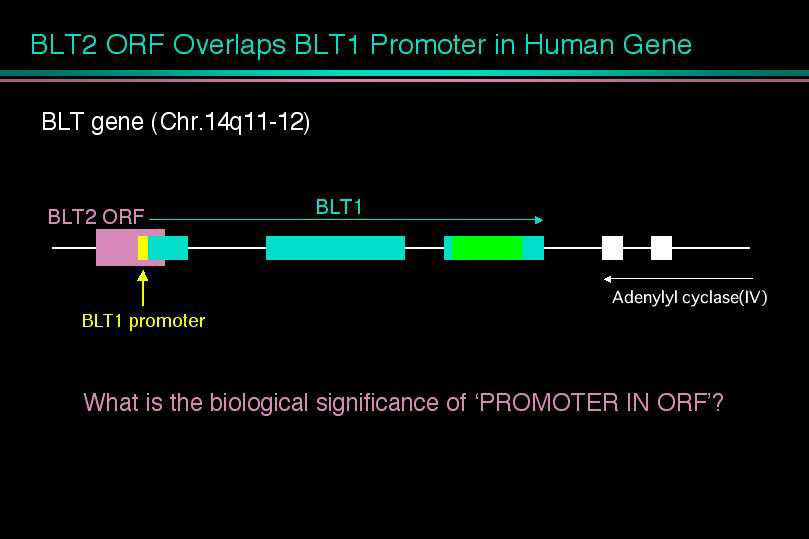
2)BLT2のタンパク質翻訳領域(ORF)は、BLT1の転写調節部位(プロモーター)と重なっており、Promoter in ORFの構造を取っていました。これは、細菌や、ウイルスで報告されている構造ですが、哺乳動物では初めて発見されたことになります。
まだここには書くことのできないおもしろいことが次々にわかってきています。私は10年間ずっとLTB4の研究をしていますが、続々とLTB4に関連した酵素や受容体が目の前に姿を現し、興味深い現象を見せてくれることに深く感動しています。LTB4には何か因縁めいたものを感じます。BLT1とBLT2の関係や、Promoter in ORFについて研究してみたい大学院生やPost Docの希望者がいらっしゃいましたら、遠慮なく御連絡下さい。清水孝雄教授か、横溝岳彦までメイルでどうぞ。
1)Yokomizo, T., Kato, K., Terawaki, K., Izumi, T., and Shimizu, T. A second leukotriene B4 receptor, BLT2: a new therapeutic target in inflammation and immunological disorders. J. Exp. Med. (2000) 192: p421-432
2)Kato, K., Yokomizo, T., Izumi, T., and Shimizu, T. Cell-specific transcriptional regulation of human leukotriene B4 receptor gene. J. Exp. Med. (2000) 192: p413-420
3)Noiri, E., T. Yokomizo, A. Nakao, T. Izumi, T. Fujita, S. Kimura, and T. Shimizu. 2000. A novel in vivo approach showing the chemotactic activity of leukotriene B4 in acute renal ischemic-reperfusion injury. Proc. Natl. Aca. Sci. U.S.A. (2000) 97:p823-828.
4)Yokomizo, T., Izumi, T., Chang, K., Takuwa, Y., and Shimizu, T. A G-protein coupled receptor for leukotriene B4 that mediates chemotaxis. Nature (1997) 387:p620-624
LTB4について知りたいという方は以下の総説を参照してください。
5)Serhan, C.N., J.Z. Haeggstrom, and C.C. Leslie. Lipid mediator networks in cell signaling: update and impact of cytokines. Faseb. J. (1996)10:1147-1158
6)Yokomizo, T., K. Masuda, K. Kato, A. Toda, T. Izumi, and T. Shimizu. Leukotriene B4 receptor. Cloning and intracellular signaling. Am. J. Respir. Crit. Care. Med. (2000) 161 S51-55
7)横溝岳彦 ロイコトリエンの代謝と受容体 細胞工学 (1998) 17 p722-729
8)Charles
N. Serhan and Stephen M. Prescott. The Scent of a Phagocyte: Advances
on Leukotriene B4 Receptors. J. Exp. Med. (2000)192:F5-F8
 横溝岳彦のホームページへ戻る
横溝岳彦のホームページへ戻る
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}